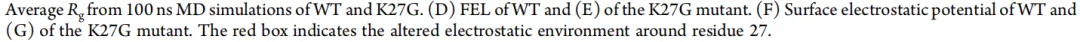2026 年 3 月 5 日,来自长沙理工大学的Ying Lv等人在Journal of Agricultural and Food Chemistry上发表了一篇题为Mining, Identification, and Engineering of DbCel5A, a Thermostable Processive Endoglucanase with Potential for One-Step Cellulose Hydrolysis的研究性论文。纤维素是储量最丰富的木质纤维素资源,其生物转化需要高效的纤维素酶。本研究通过数据库挖掘,鉴定出源自网球菌科细菌的一种高度稳定的持续作用内切葡聚糖酶DbCel5A,并将其在大肠杆菌中成功表达。DbCel5A 表现出卓越的热稳定性,其熔化温度(Tₘ)为 79.9 °C,在 75 °C 下的半衰期约为 30 小时。高耐盐性是其一个显著特点。以葡萄糖和纤维二糖为主的水解产物谱进一步凸显了 DbCel5A 在一步法糖化中的巨大潜力。采用人工智能辅助的蛋白质工程改造,获得了最优突变体 K27G,其催化效率提高了 2.5 倍,同时具有更高的底物亲和力和增强的热稳定性(Tₘ = 81.9 °C)。分子动力学模拟揭示了其深层机制,并且通过对甘蔗渣的有效降解证实了其实际应用性。总体而言,DbCel5A 是一种极具前景的生物催化剂,并为高性能纤维素酶的发现与改造提供了一种有效策略。
Introduction
全球对可持续能源和生物基产品日益增长的需求,已将木质纤维素生物质转化为可发酵糖推至生物技术研究的前沿。作为地球上最丰富的有机聚合物,纤维素代表着巨大的可再生碳资源库。其高效降解依赖于纤维素酶的协同作用。纤维素酶是糖苷水解酶家族中的一个多组分酶系统,主要包括内切葡聚糖酶(EG;EC 3.2.1.4)、纤维二糖水解酶(CBH;EC 3.2.1.91)和β-葡萄糖苷酶(BG;EC 3.2.1.21)。这些酶协调作用:内切葡聚糖酶随机切割纤维素无定形区域的糖苷键,纤维二糖水解酶从结晶链末端持续释放纤维二糖,β-葡萄糖苷酶随后将纤维二糖水解为葡萄糖。除这些核心组分外,包括膨胀因子、纤维二糖脱氢酶和裂解性多糖单加氧酶在内的几种辅助酶也能增强纤维素的可及性,并显著提高纤维素酶系统的整体水解效率。凭借这种催化多功能性,纤维素酶已广泛应用于生物燃料生产、纺织加工、食品改良和动物饲料优化等领域。
天然纤维素酶的工业应用常常因其热稳定性不足而受限。嗜热酶在高温下仍保持高催化活性,是用于木质纤维素转化的有前景的生物催化剂。目前获取耐热纤维素酶的方法包括:从嗜热环境中进行宏基因组挖掘,以及通过蛋白质设计或人工智能引导的优化对现有酶进行稳定性改造。Trachtmann等人通过生物信息学分析,从上巴尔卡尔地下深层热泉的环境微生物宏基因组中发现了一个新型纤维素酶基因,并获得了重组内切葡聚糖酶Cel7465,该酶在80°C下半衰期约为1小时。此外,基于源自枯草芽孢杆菌 BS-5的GH5家族酶EG5C-1,结合聚类分析和贪心算法预测得到一个突变体(M8),该突变体在60°C孵育2小时后,其残余活性比野生型高出约20%。
传统上,纤维二糖水解酶被认为是唯一能够持续性降解纤维素的纤维素酶。后续研究发现,某些内切葡聚糖酶也具有持续性水解纤维素的能力,展现出内切/外切双重功能。自1993年首次报道以来,这些持续性内切葡聚糖酶已获得广泛关注。它们不仅能切割纤维素无定形区域内的内部糖苷键,还能通过沿纤维素链持续滑动来持续产生可溶性还原糖,与传统非持续性内切葡聚糖酶相比,能释放出更高产量的可溶性寡糖。这种独特的机制使其能够彻底解构纤维素,成为生物质转化领域颇具前景的生物催化剂。尽管如此,已被表征的持续性内切葡聚糖酶仍然有限(表1),且很少有能满足苛刻工业应用所需的卓越热稳定性。例如,源自喜马拉雅芽孢杆菌 PCH94、巴塞罗那类芽孢杆菌及其他相关细菌种类的几种持续性内切葡聚糖酶表现出有限的热稳定性,在50-70°C的中高温下仅能维持约1小时甚至仅几十分钟的催化活性。类似地,来自海栖热袍菌的TmCel5A虽然是嗜热的,但其热稳定性仍有相当大的提升空间。这一差距凸显了发现具有更强稳定性和活性的新型持续性内切葡聚糖酶的必要性。
本研究旨在通过数据库基因挖掘和机器学习指导的诱变,构建一种新型且高度耐热的持续性内切葡聚糖酶。以海栖热袍菌 MSB8的GH5家族持续性内切葡聚糖酶基因为模板。通过在NCBI数据库中进行基因挖掘,从网球菌科细菌的GH5家族中鉴定出一种天然的持续性内切葡聚糖酶(DbCel5A)。该酶在大肠杆菌中成功表达和纯化,并对其生化特性和水解产物谱进行了全面表征。在第二阶段,采用机器学习预测和筛选氨基酸突变位点,旨在开发能够协同增强热稳定性和催化效率的突变体。这项工作为生物质资源的高值化转化提供了理论基础,并推动了持续性内切葡聚糖酶的工业应用。
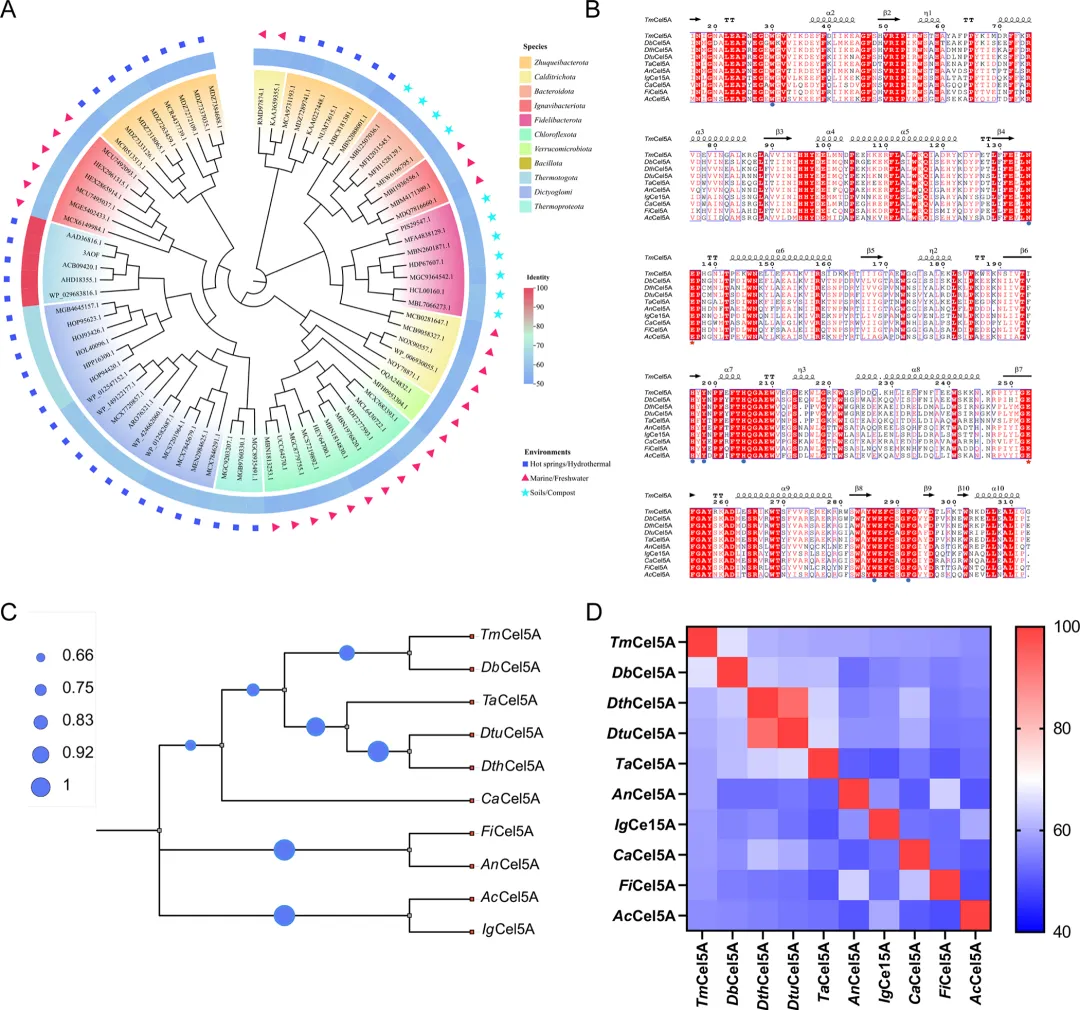
为满足当前工业对高度耐热持续性内切葡聚糖酶的需求,通过文献调研(表1)与基于数据库的筛选策略相结合,对目标酶进行了序列挖掘与分析。为了准确高效地识别耐热持续性内切葡聚糖酶,以海栖热袍菌 MSB8来源的耐热酶TmCel5A为模板,选取与TmCel5A氨基酸序列一致性为50−70%的序列作为潜在的功能候选者。使用MEGA 11对所选序列进行多序列比对,随后根据物种来源和生态环境构建系统发育树(图1A)。通过系统发育树的根位和枝长推断这些序列的分类学来源。分析结果表明,所选序列主要源于朱鹊杆菌门和嗜热网球菌,并在伊格纳维菌门细菌、厌氧绳菌纲和Fidelibacterota门中也鉴定到了其他代表性序列。
随后,采用ProtParam分析,选取预测理论半衰期大于10小时且催化活性位点与TmCel5A一致的氨基酸序列,进行进一步的性质预测和筛选。如序列比对结果(图1B)所示,选定的九个氨基酸序列均具有高度保守的催化残基(Glu156和Glu273)和底物结合位点(Trp50、Asn155、His196、Tyr198、His225、Trp306和Phe312)。这些保守特征表明,所选候选蛋白与模板酶TmCel5A具有相似的催化特性,并可能具备持续性催化潜力。来自嗜热微生物的酶在长期高温条件下进化,通常表现出更强的结构稳定性和热适应性,因而更有可能兼具耐热性和高催化效率⁵⁰。鉴于本研究旨在鉴定一种具有优异耐热性的持续性内切葡聚糖酶,特别关注了源自嗜热类群(如网球菌纲和热袍菌门)的候选序列。通过这一多步筛选策略,最终选择了与模板酶TmCel5A亲缘关系最近的来自网球菌科细菌的糖苷水解酶(DbCel5A,GenBank登录号:HOP94420.1)进行深入研究。序列相似性的可视化结果如图1D所示。
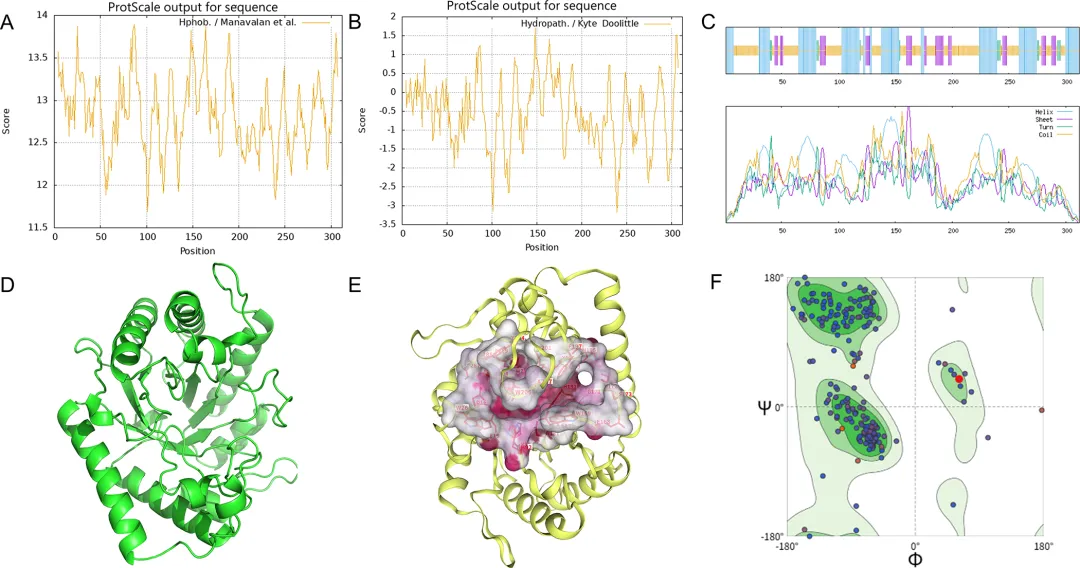
2. 重组 DbCel5A 的生物信息学表征、表达与纯化DbCel5A 基因编码一个由 312 个氨基酸残基组成的多肽,理论分子量为 36.7 kDa。利用 Expasy 和 ProteinTools 网站上的工具预测了 DbCel5A 的生物学特性,结果如图 2 所示。如图 2A 所示,DbCel5A 中每个氨基酸残基的亲水性分析得出平均疏水值为 -0.543,表明该蛋白质总体上是亲水的。图 2B 描绘了 DbCel5A 的表面环境,显示出显著的整体疏水性,并伴有局部分布的明显疏水斑块。预测的 DbCel5A 二级结构(图 2C)主要由 α-螺旋(37.5%)、延伸 β-链(14.1%)和无规则卷曲(43.9%)组成。以 TmCel5A 的 3D 结构为模板,构建了 DbCel5A 的三级结构模型(图 2D)。使用 CB-Dock2 预测了 3D 模型中的潜在催化活性位点和底物结合口袋(图 2E),鉴定出的催化残基与多序列比对揭示的保守氨基酸一致。根据拉马钱德兰图分析,91.09% 的残基位于允许区域,表明该构象合理且拟合良好(图 2F)。
使用 ProteinTools 对 3D 模型进行了进一步的结构分析,评估了疏水簇、氢键网络、盐桥和残基间距离。在催化位点周围鉴定出七个疏水簇,其中四个紧密堆积在活性口袋附近。此外,检测到 16 个氢键和 11 个盐桥,它们都有助于催化位点的结构完整性和功能稳定性。预测的残基间距离与 3D 结构模型中观察到的距离很好地吻合。
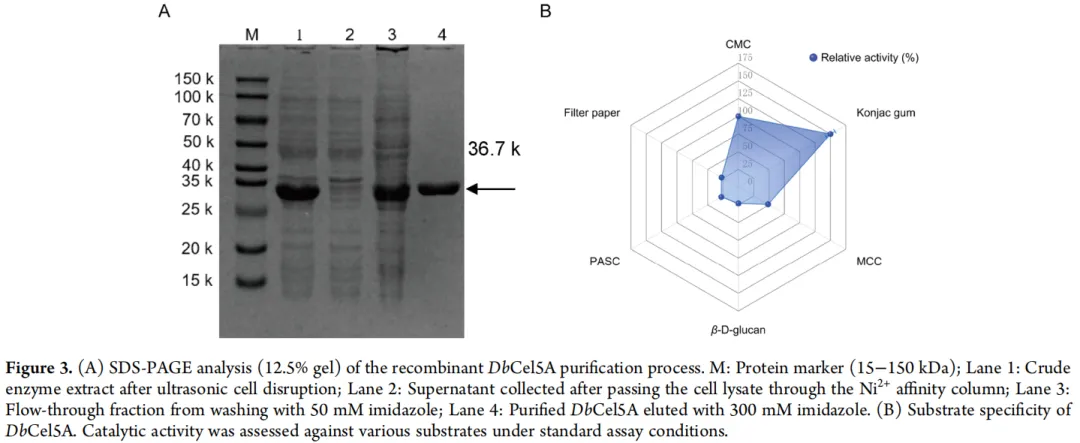
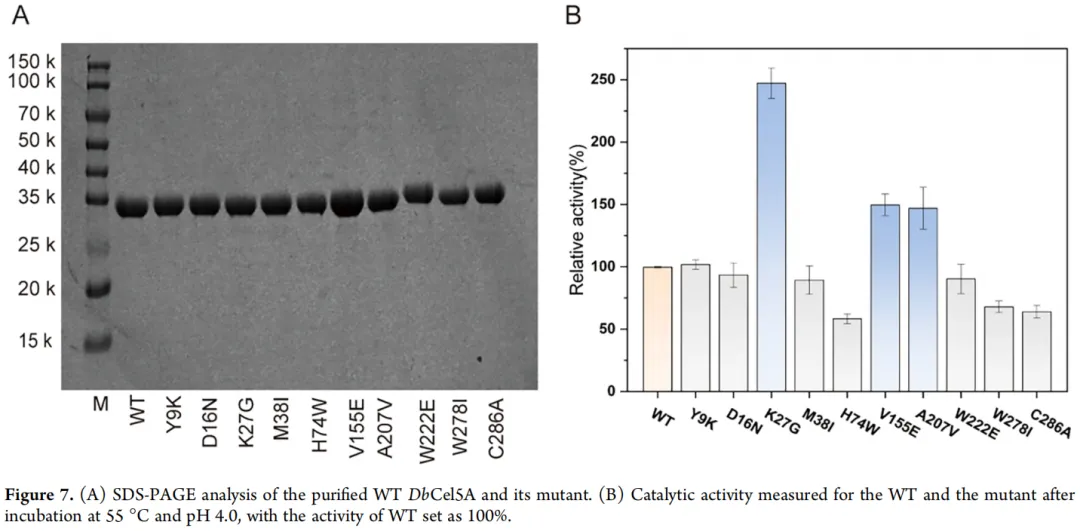
使用 pET-22b 载体在大肠杆菌 BL21(DE3) 中异源表达了编码 DbCel5A 的基因。通过镍亲和层析纯化重组蛋白。纯化蛋白的 SDS-PAGE 分析显示出一条与目标蛋白相对应的、分子量约为 35 kDa 的清晰单一条带(图 3),证实了 DbCel5A 的可溶性和高效表达。在标准反应条件下测定了 DbCel5A 的底物特异性。如图 3B 所示,该酶对可溶性底物(包括 CMC 和葡甘露聚糖)表现出高催化活性,揭示了其特征性的内切葡聚糖酶活性及其降解半纤维素(葡甘露聚糖)的潜力。相反,对不溶性底物(如 MCC 和滤纸)检测到的活性显著较低。这种底物特异性与持续性内切葡聚糖酶 p4818Cel5_2A和 H1AD14的催化行为相当,显示出对可溶性多糖的强大活性。
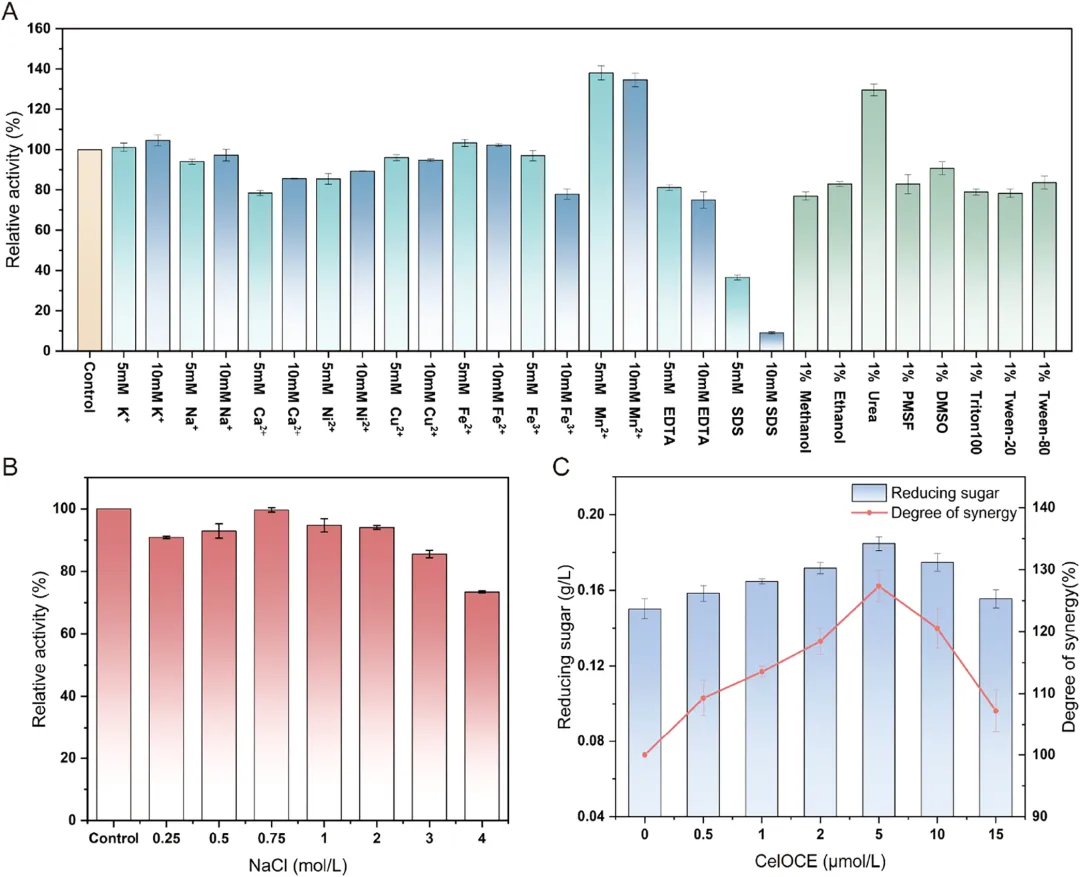
实验以 1% CMC 为底物,对 DbCel5A 的酶学性质进行表征。研究了不同温度、pH 值、金属离子、有机试剂、表面活性剂以及不同浓度氯化钠对 DbCel5A 活性的影响。结果如图 4 和图 5 所示。
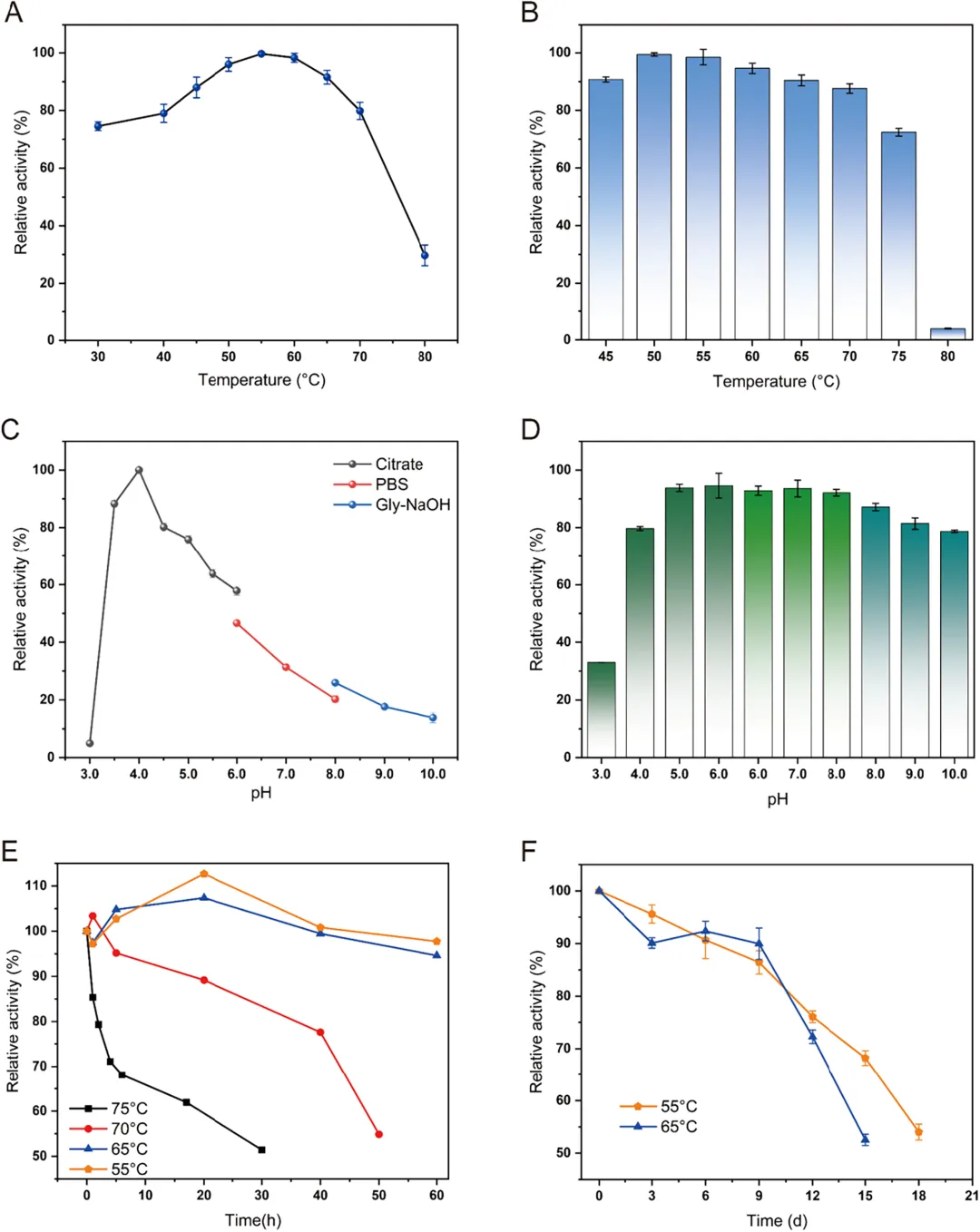
图 4A 展示了 DbCel5A 在 30 至 80 °C 温度范围内的活性变化。实验结果表明,DbCel5A 在 55 °C 时表现出最高酶活性,且随着温度升高,活性呈现先升高后降低的趋势。该酶具有较宽的反应温度范围,在 45 至 70 °C 之间保持高催化活性(20 分钟内超过 80%)。然而,在 80 °C 时,酶活性显著下降,这可能是由于高温条件下催化结构失稳所致。此外,DbCel5A 表现出很强的热稳定性。图 4B 评估了 DbCel5A 的热稳定性,测量了酶在 45 至 80 °C 温度下孵育 2 小时后的残余酶活性。该酶在低于 75 °C 的温度下能保持超过 70% 的活性。与温度变化相比,DbCel5A 的催化效率受 pH 值波动的影响更为显著。图 4C 数据显示,DbCel5A 的最适反应 pH 为 4.0,其催化活性依赖于弱酸性条件。该酶在强酸性(pH < 3)和碱性条件(pH > 8)下会显著失活;然而,在 pH 3.5 至 5.0 范围内,其相对酶活性保持在 70% 以上。当 pH 升至 6.0 和 8.0 时,酶活性分别急剧下降至约 50% 和 20%。尽管如此,与其对 pH 波动的显著敏感性相反,该酶在宽 pH 范围内表现出优异的结构稳定性。如图 4D 所示,即使在中性和碱性缓冲液中孵育 2 小时后,其残余酶活性损失极小。这些结果表明,DbCel5A 是一种催化活性相对偏酸性,但在结构稳定性方面对宽 pH 范围具有良好耐受性的酶。
由于初步实验中观察到 DbCel5A 具有很强的耐热性,本研究进一步考察了该酶在最适温度及以上的半衰期(图 4E)。结果显示,该酶在 70 °C 下半衰期约为 30 小时,在 75 °C 下半衰期约为 50 小时。相反,在 55 °C 和 65 °C 下孵育 60 小时后,酶活性没有显著下降,而是呈现轻微增加后缓慢降低的趋势。将 DbCel5A 在 55 °C 和 65 °C 下进一步孵育发现,活性在 9 天后开始下降(图 4F),并分别在 15 天和 18 天后最终降至初始活性的 50% 左右。这些发现表明 DbCel5A 具有卓越的结构稳定性。相比之下,大多数报道的嗜热内切葡聚糖酶在高温下的稳定性显著较低。例如,来自海栖热袍菌 MSB8 的模板酶 TmCel5A 在 70 °C 下仅 2 小时就失去 40% 的活性,而新发现的酶表现出更强的热稳定性。在现有研究中,EG5C-1 被认为是催化活性最高、改造最广泛的内切葡聚糖酶;然而,它在 70 °C 下 2 小时后仅保留 30% 的稳定性。相比之下,DbCel5A 在高温下表现出更优的热稳定性。此外,与表 1 中列出的其他持续性内切葡聚糖酶相比,DbCel5A 在高温下表现出最佳的热稳定性。DbCel5A 卓越的热性能表明其具有巨大的工业应用潜力。随后,DSF 实验确定 DbCel5A 的Tₘ 为 79.9 °C,这支持了 80 °C 时酶活性显著变化是由于其三维结构失稳所致的结论。DbCel5A 的高Tₘ 表明该蛋白质能在高温下保持结构和功能的稳定性,这与实验观察结果一致。这种稳健的蛋白质结构可能是分子内氢键和疏水相互作用协同效应的结果。
为了进一步阐明 DbCel5A 优异热稳定性和持续催化能力的结构基础,与先前报道的两个单模块持续性内切葡聚糖酶 TmCel5A(PDB: 3AMC)和 EG5C-1(模板 PDB: 3PZT)进行了系统的结构比较。这两种酶都属于 GH5 家族。结构比对结果显示,三种酶均具有 GH5 家族典型的 (β/α)₈ TIM-桶状折叠结构,表明催化骨架高度保守。然而,在几个结构特征上观察到了显著差异。分子内相互作用分析表明,DbCel5A 在催化桶区域周围形成更多的疏水簇,这可能有助于增强整体结构刚性。此外,与其他 GH5 酶相比,其疏水核心显示出更紧密的堆积排列,表明内部结构稳定性更高。此外,DbCel5A 在其环区具有更高比例的脯氨酸残基,这有助于限制主链运动,从而增强热稳定性。与表 1 中列出的其他持续性内切葡聚糖酶相比,DbCel5A 在高温下也表现出最佳的热稳定性。DbCel5A 卓越的热性能表明其具有巨大的工业应用潜力。
评估了不同浓度金属离子、有机试剂和表面活性剂对 DbCel5A 活性的影响(图 5A)。在所测试的金属离子中,Mn²⁺ 表现出最显著的刺激作用,5 mM Mn²⁺ 将酶活性提高至初始水平的 138.0%。相比之下,其他金属离子仅引起活性的微小波动,表明 DbCel5A 具有相当大的金属耐受性。与金属离子相比,有机试剂和表面活性剂对 DbCel5A 的影响更显著。除尿素外,所有其他化学试剂均导致轻微抑制,而 1% 尿素将催化活性提高至未添加化学试剂对照组的 130.0%。在所有测试因素中,阴离子表面活性剂 SDS 对 DbCel5A 的抑制效应最强。与来自地衣芽孢杆菌的高度耐热持续性内切葡聚糖酶 H1AD14⁵¹ 相比,DbCel5A 在不同金属离子存在下表现出更高的稳定性。
在生物燃料生产、造纸及其他工业应用中,纤维素酶的耐盐性是决定操作稳定性的关键特性。普遍认为,在 1−3 M NaCl 存在下保持 ≥ 70% 活性的纤维素酶具有理想的耐盐性。存在少数特殊的极端嗜盐纤维素酶,即使在超过 3 M NaCl 的浓度下也表现出显著活性;然而,这类酶相对罕见,且主要限于实验室研究。本研究在高盐浓度下系统评估了 DbCel5A 的耐盐性,如图 5B 所示。结果表明,DbCel5A 在 0.75 M NaCl 中的催化活性与无盐条件下几乎相同;在高盐条件下,DbCel5A 在 3 M 和 4 M NaCl 中分别保持约 86% 和 73% 的活性,表现出很强的耐盐性。Trivedi报道,从石莼中分离的一种耐盐纤维素酶在 2.5 M NaCl 中保持 70% 的活性,但在 3.5 M NaCl 中降至 18%。即使是来自海洋芽孢杆菌的 Bc22Cel,在 1.5 M NaCl 中孵育 1 小时后仍保持 70% 的残余活性,也被认为具有耐盐性。总体而言,DbCel5A 的耐盐性超过了大多数已报道的非嗜盐细菌来源的纤维素酶,凸显了其作为盐环境中生物催化剂的潜力。
4. CelOCE 与 DbCel5A 协同增强结晶纤维素降解CelOCE 是最近通过宏基因组分析发现的一种来源于 AA18 家族的新型裂解性多糖单加氧酶,其具有强大的氧化切割纤维素的能力。这种能力使其能够更有效地破坏纤维素链,使纤维素的结晶区更易于被水解酶降解。当使用微晶纤维素作为底物时,添加 CelOCE 可增强 DbCel5A 对结晶纤维素的降解能力。协同效应在 CelOCE 浓度为 5 μM 时最为显著,与单独使用 DbCel5A 相比,还原糖产量提高了 28%。这表明 DbCel5A 具备与氧化切割酶协同有效降解结晶纤维素的能力。然而,当 CelOCE 浓度超过 5 μM 时,协同效应逐渐减弱;在 15 μM 时,还原糖产量仅为 5 μM 时的 84%(图 5C)。这种下降可能归因于过量 CelOCE 的氧化位点与纤维素酶催化位点之间的竞争性吸附,导致协同作用降低。因此,选择两种酶的最适比例可以实现更有效的纤维素降解。
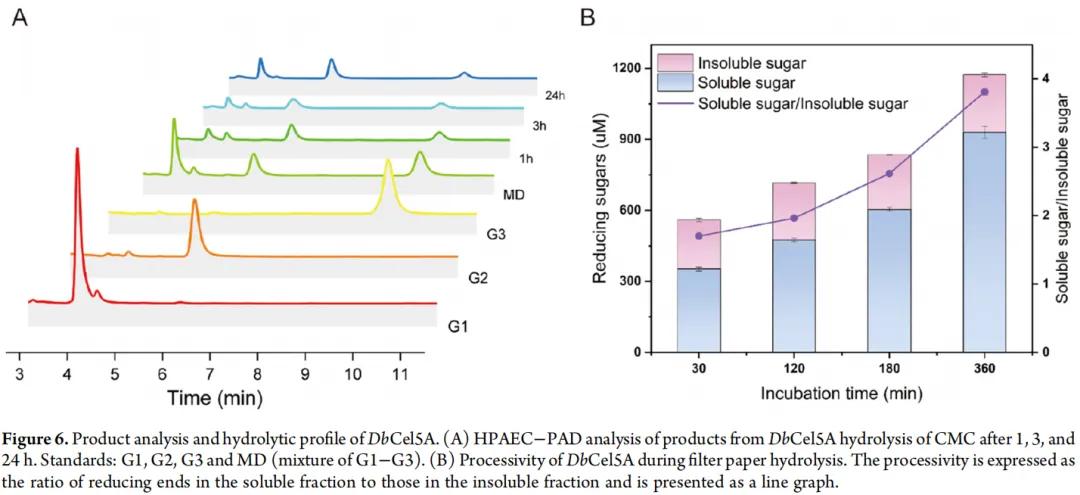
以 1% CMC 为底物,通过分析反应 1、3 和 24 小时后水解产物的组成,研究了 DbCel5A 的催化模式。图 6A 显示了 DbCel5A 产生的水解产物的 HPAEC-PAD 分析结果。结果表明,DbCel5A 在水解 CMC 过程中主要产生小分子寡糖(G1、G2 和 G3)。随着反应进行至 24 小时,可以观察到葡萄糖和纤维二糖明显增加,同时纤维三糖显著减少,表明大多数 G3 分子被进一步水解为 G1 和 G2。这种产物模式偏离了典型内切葡聚糖酶的传统定义,后者通常报道以纤维二糖为主要产物,并伴有少量其他短链寡糖。相比之下,DbCel5A 对纤维素底物的水解更为彻底,以 G1 和 G2 为主要产物。如表 1 所示,大多数已报道的其他来源的持续性内切葡聚糖酶主要产生 G2 和 G3,仅产生痕量的 G1。因此,DbCel5A 似乎表现出一种独特的催化特征,即产生更彻底的水解产物,这显示了其通过单一酶实现一步法完全水解纤维素的潜力。
为了验证这些观察结果,还进行了高效液相色谱分析。高效液相色谱结果与 HPAEC-PAD 获得的结果完全一致,表明随着反应进行,G1 和 G2 仍然是主要产物。这种水解模式表明,DbCel5A 可能能够以近乎完全的方式独立地将纤维素降解为小分子糖,从而实现纤维素到葡萄糖的高效转化。
6. DbCel5A 持续作用能力的验证
具有内切/外切双重活性的内切葡聚糖酶的持续作用能力,可以通过其水解定性滤纸产生的可溶性还原糖与不溶性还原糖的比值来量化。S/I 比值在 3 到 8 之间表明具有持续作用特性。如图 6B 所示,DbCel5A 水解 1% Whatman 滤纸的 S/I 值在 6 小时后增加至 3.8,反映了其显著的持续水解能力。Wang 等人报道的持续性内切葡聚糖酶 EG1-Δ19 突变体也观察到了类似现象,其在滤纸上的 S/I 比值从 1.6 增加到 4.7。结合水解产物分析,这些结果证实 DbCel5A 是一种持续性内切葡聚糖酶。
基于前述发现,DbCel5A 被鉴定为一种高度耐热但催化效率较低的持续性内切葡聚糖酶。由于其催化效率有限,DbCel5A 的工业应用仍面临挑战。VenusMutHub是一个新开发的预测模型,旨在评估蛋白质点突变的效果。它利用蛋白质结构语言模型来评估点突变如何影响不同蛋白质的活性、稳定性及其他关键功能特性。该模型通过整合来自已发表文献和公共数据库的 527 种蛋白质的 905 个数据集构建而成。使用 ColabFold 进行数据处理,并系统评估了 23 种不同的计算模型。该算法框架能够精确定量突变诱导的功能变化,为蛋白质工程和疾病相关突变的机制研究提供了强大的工具。
因此,为获得催化活性增强的突变体,本研究采用 VenusMutHub AI 算法模型,基于突变成功概率预测并筛选了最有可能提高 DbCel5A 催化性能的 10 个突变位点(Y9K、D16N、K27G、M38I、H74W、V155E、A207V、W222E、W278I 和 C286A)。成功构建了相应的突变体,并进行了表达和纯化(图 7A)。其中,三个突变体 K27G、V155E 和 A207V 与野生型酶相比表现出显著提高的催化活性,K27G 显示出最显著的 2.5 倍提升(图 7B)。此外,K27G 突变增加了酶对 CMC 底物的亲和力,将Kₘ从 13.42 mg/mL 降低至 11.30 mg/mL,并将kcₐₜ/Kₘ 比值提高了 2.43 倍(表 2)。热稳定性评估显示,K27G、V155E 和 A207V 突变体在 70 °C 孵育 2 小时后,分别保留了初始活性的 94%、77% 和 82%。值得注意的是,K27G 突变体的Tₘ 也略有增加,从 79.9 °C(野生型)升至 81.9 °C。总之,这些结果表明,第 27 位的甘氨酸取代协同增强了 DbCel5A 的催化效率和热稳定性,为相关酶的改造提供了有价值的指导。
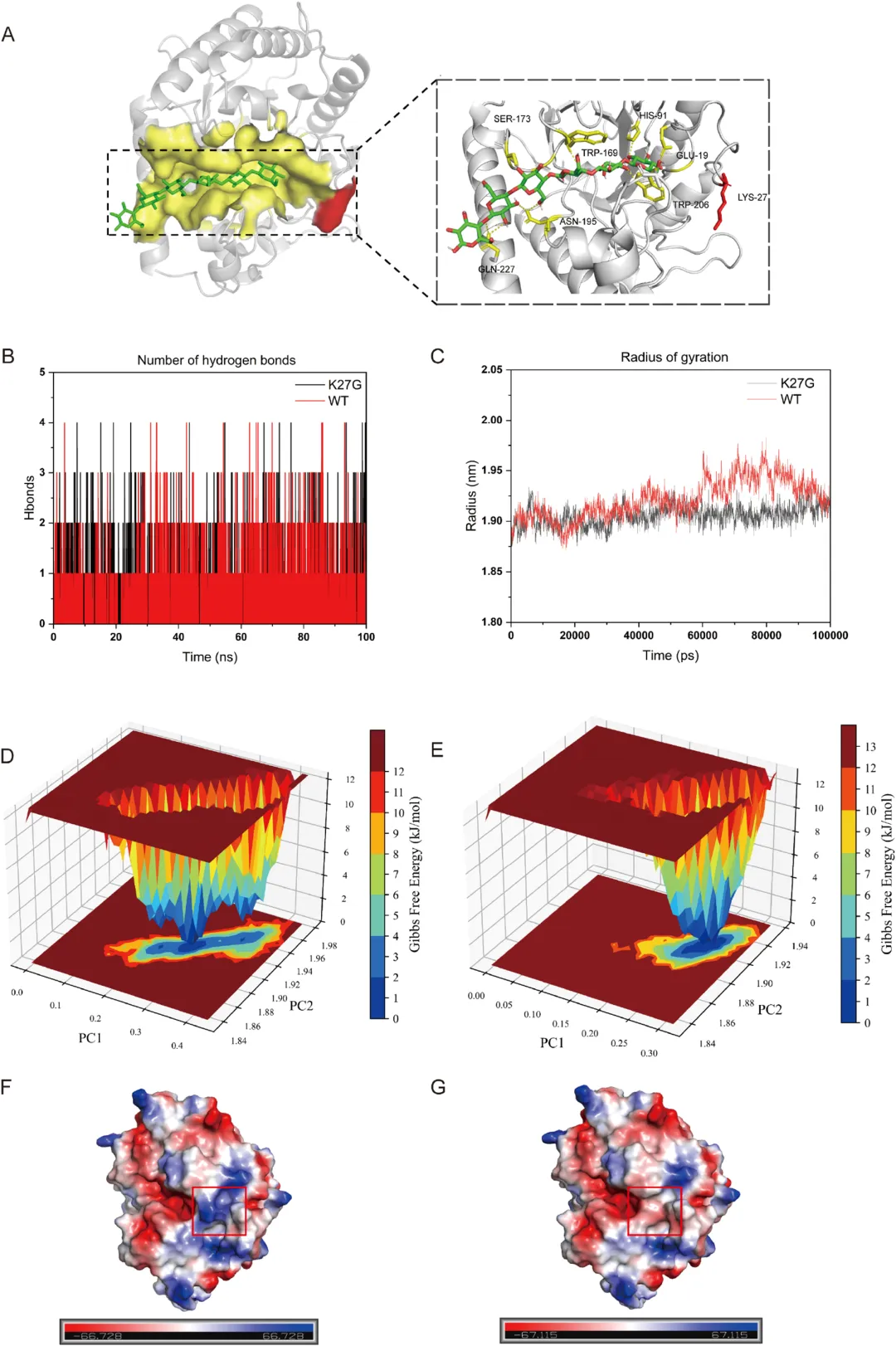
为了研究 K27G 突变体酶学性质增强的结构基础,对野生型和 K27G 突变体与纤维六糖进行了分子对接和 100 ns 的分子动力学模拟。图 8 所示的结果揭示了 K27G 酶学性质改善的分子机制。图 8A 展示了 DbCel5A 与纤维六糖的结合模式,并突出了配体 3.5 Å 范围内的氨基酸残基。在 100 ns 分子动力学轨迹上的氢键分析表明,K27G 形成的平均氢键数略高于野生型(0.57 vs 0.52),并且在整个模拟过程中表现出更高的氢键形成事件频率,表明局部结构稳定性有适度增强(图 8B)。通过计算回转半径评估蛋白质结构致密性。在模拟的前 50 ns 之后,K27G 始终保持比野生型更低的回转半径(图 8C)。减小的波动和略低的回转半径表明 K27G 采用了更致密和稳定的构象,这可能有利于底物结合并有助于提高催化效率。自由能形貌图分析进一步揭示了野生型和 K27G 之间不同的构象分布(图 8D、E)。突变体沿主成分表现出更集中、更深的能量盆地,对应于一个主要的低能态,而野生型则表现出更宽、更拉长的能量盆地,表明更高的构象柔性。用小的中性甘氨酸替换长链带正电的赖氨酸,将局部静电势从正变为微负,并减弱了周围的电荷效应(图 8F、G)。这可能减轻静电排斥,并为活性位点内的底物重定位提供空间柔性。总之,这些结果表明 K27G 突变通过稳定低能态增强了构象稳定性,这可能有助于其催化性能的提高。
甘蔗渣是一种常见的高结晶度木质纤维素生物质资源。以甘蔗渣为底物,对酶学性质改善的工程突变体 K27G 的降解性能进行了评估,以考察其在实际木质纤维素生物转化中的应用潜力。XRD 结果表明,未经处理的甘蔗渣(CK 组)在 2θ = 15°、22° 和 34° 处显示出特征衍射峰,这对应于天然纤维素的典型晶体结构。经 K27G 处理 12 小时后,衍射峰强度显著降低,表明结晶度降低,有序纤维素结构被破坏。图 S5B 展示了 K27G 水解甘蔗渣产物的高效液相色谱分析结果。结果显示,纤维二糖是唯一可检测到的产物。这些发现表明,K27G 能够破坏甘蔗渣的晶体结构,并具有相当大的工业应用潜力。
总之,通过数据库挖掘和各种生物信息学分析,从网球菌科细菌中鉴定出一个来源于 GH5 家族的新型持续性内切葡聚糖酶基因 DbCel5A。该酶表现出卓越的热稳定性,在高温下长时间孵育后仍保持显著活性,并具有高的Tₘ 值。DbCel5A 对盐和化学试剂表现出很强的耐受性,凸显了其稳健性。通过与裂解性多糖单加氧酶的协同作用,其催化活性得到进一步提升。水解产物以葡萄糖和纤维二糖为主,表明其具有彻底的一步法纤维素水解能力。此外,人工智能辅助的蛋白质工程产生了活性和热稳定性均提高的 K27G 突变体,并分析了 K27G 性能改善的分子机制。最后,XRD 和高效液相色谱分析证实了 K27G 对实际底物甘蔗渣的有效水解,为生物质资源的高值化利用提供了新的技术支撑。基于 DbCel5A 的稳定性,未来的研究可采用人工智能辅助设计进行定向进化,使其适应不同的工业环境(例如特定的 pH、离子强度),并研究其催化机制,从而促进更高效、更具成本效益的生物质转化。